|
Performing a hunt by interaction
mating
Authors: Mikhail G. Kolonin and Russell L. Finley Jr.
Affiliation: Center for Molecular Medicine and Genetics
Wayne State University School of Medicine
540 East Canfield Ave.
Detroit, Michigan 48201
Abstract
When more than one bait will be used to screen a single library,
significant time and resources can be saved by performing the
interactor hunt by interaction mating. In this protocol one strain is
transformed with library DNA and the transformants are collected and
frozen in aliquots. For each interactor hunt, an aliquot of this
frozen "pretransformed library strain" is thawed and mixed with an
aliquot of a bait strain transformed with the bait expression
plasmid. Overnight incubation of the mixture on a YPD plate results
in mating - i.e., cells of one strain fuse with cells of the other
strain to form diploids. The diploids are then exposed to galactose
to induce expression of the library encoded proteins, and interactors
are selected in the same manner as in the standard hunt protocols.
The advantage to this approach is that it requires only one
high-efficiency library transformation for multiple hunts with
different baits. It is also useful for bait proteins that are
somewhat toxic to yeast; yeast expressing toxic baits can be
difficult to transform with the library DNA.
Back to Top
Background and Introduction
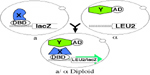
An alternate way of conducting an interactor hunt is to mate a
strain that expresses the bait protein with a strain pretransformed
with the library DNA, and screen the resulting diploid cells for
interactors (Bendixen et al., 1994; Finley and Brent, 1994). This
"interaction mating" approach can be used for any interactor hunt,
and is particularly useful in three special cases. The first case is
when more than one bait will be used to screen a single library.
Interaction mating allows several interactor hunts with different
baits to be conducted using a single high-efficiency yeast
transformation with library DNA. This can be a considerable savings
of time a resources, since the library transformation is one of the
most challenging tasks in an interactor hunt . Moreover, some yeast
strains pretransformed with libraries are becoming commercially
available, which may eliminate altogether the need to conduct a
high-efficiency library transformation for some researchers. The
second case is when a constitutively expressed bait interferes with
yeast viability. For such baits, performing a hunt by interaction
mating avoids the difficulty often encountered when trying to achieve
high-efficiency transformation of a strain expressing a toxic bait.
Moreover, the actual selection for interactors will be conducted in
diploid yeast, which are more vigorous than haploid yeast and
generally can better tolerate expression of toxic proteins. The third
case is when a bait cannot be used in a traditional interactor hunt
using haploid yeast strains because it activates transcription of
even the least sensitive reporters. In diploids the reporters are
less sensitive to transcription activation than they are in haploids.
Thus, the interaction mating hunt provides an additional method to
reduce background from transactivating baits.
In the protocol described below, the library DNA is used to
transform a strain with a LEU2 reporter. This pretransformed
library strain is then frozen in many aliquots, which can each be
thawed and used for individual interactor hunts. The bait is
expressed in a strain of mating type opposite to that of the
pretransformed library strain, and also bearing the lacZ reporter. A
hunt is conducted by mixing the pretransformed library strain with
the bait strain and allowing diploids to form on YPD medium
overnight. The diploids are then incubated in galactose to induce
expression of the library encoded proteins, and screened for
interactors in the same manor as in a traditional haploid interactor
hunt.
NOTE: Strain combinations other than those described below can
also be used in an interaction mating hunt. The key to choosing the
strains is to ensure that the bait and prey strains are of opposite
mating types and that both have auxotrophies to allow selection for
the appropriate plasmids and reporter genes. Also, once the bait
plasmid and lacZ reporter plasmid have been introduced into the bait
strain, and the library plasmids have been introduced into the
library strain, the resulting bait strain and library strain must
each have auxotrophies that can be complemented by the other, so that
diploids can be selected.
The interaction mating protocol presented here is based on the
LexA version of the two-hybrid system (Fields and Song, 1989)
developed in Roger Brent's lab (e.g., Gyuris et al., 1993). Go here
to find links to background information and other protocols for the
Brent lab system. Briefly, the essential features of the Brent lab
system are:
- The reporter is a LEU2 gene with upstream LexA binding sites
or operators (LexAop-LEU2) integrated into the yeast genome in
strains like EGY48 (Estojak et al., 1995) or a derivative of
EGY48, RFY231 (Kolonin and Finley, 1998). Different strains with
different numbers of LexA, and hence different levels of
sensitivity, are available (Estojak et al., 1995).
- The interaction assay is usually conducted in the presence of
an additional lacZ reporter. The LexAop-lacZ reporter usually
resides on a multicopy (2µm) plasmid bearing a URA3 marker,
such as pSH18-34. Again, various lacZ reporters with different
numbers of upstream LexA operators exist. Strains with the entire
lacZ plasmid integrated into the ura3 locus also are available;
these lacZ reporters are the least sensitive as they are in single
copy.
- The LexA fusion or bait is expressed from a multicopy
(2µm) bait plasmid bearing a HIS3 marker, such as pEG202
(Estojak et al., 1995). The LexA bait is constitutively expressed
from the yeast ADH1 promoter. Version of the bait plasmid also
exist with a nuclear localization signal fused to LexA, or with
the ADH promoter replaced by the GAL1 promoter for conditional
expression of bait.
- The library or "prey" plasmid (i.e., expressing the activation
domain (AD) -fused protein that may interact with the bait) is a
multicopy (2µm) plasmid bearing the TRP1 marker, such as
pJG4-5 (Gyuris et al., 1993). The AD-fused protein is expressed
from the yeast GAL1 promoter - on in galactose but off in
glucose.
Back to Top
Protocol (20
steps)
Materials
Yeast Strains:
- Bait strains: Either RFY206 (Finley and Brent, 1994), YPH499
(Sikorski and Hieter, 1989; ATCC#6625), or an equivalent
MATa strain with auxotrophic markers ura3, trp1, his3, and
leu2.
Library strains: Either RFY231 (Kolonin and Finley, 1998),
EGY48 (Estojak et al., 1995), or other MATalpha strain with
LexAop-LEU2 reporter and auxotrophic markers ura3, trp1, his3, and
leu2.
Media and Solutions
Construct the bait
strain
The bait strain will be a MATa yeast strain (mating type
opposite of RFY231) containing a lacZ reporter plasmid like pSH18-34
and the bait-expressing plasmid, pBait.
1. Construct the bait plasmid (pBait) to express the LexA
fused bait protein, for example using the HIS3 2µm plasmid
pEG202 (Estojak et al., 1995).
2. Co-transform the MATa yeast strain (e.g., either
RFY206 or YPH499) with pBait and a lacZ reporter, such as the very
sensitive pSH18-34, using the lithium
acetate method. Select transformants on Glu/CM-Ura,-His plates.
Incubate plates at 30oC for 3-4 days until colonies form.
Combine 3 colonies for all future tests and for the mating hunt.
- The bait strain (RFY206/pSH18-34/pBait or
YPH499/pSH18-34/pBait) can be tested by immunoblotting to ensure
that the bait protein is expressed.. Synthesis and nuclear
localization of the bait protein can also be tested by the
repression assay, as described in previous protocols.
3. (optional) Assay lacZ gene activation in the bait
strain, for example, by plating to Glu/CM-Ura,-His X-Gal plates.
- If the bait activates the lacZ reporter, a less sensitive lacZ
reporter plasmid, or an integrated version of the lacZ reporter
should be tried. A bait that strongly activates the lacZ reporters
usually cannot be used in a hunt based on selection of interactors
with the LEU2 reporter because the LEU2 reporters are more
sensitive than even the most sensitive lacZ reporters. However,
both reporters are less sensitive to activation by a bait in
diploid cells, as compared to haploid cells. Thus, a more
important test of the transactivation potential of a bait is to
test the leucine requirement of diploid cells expressing it, as
described in steps 6 - 19 of this protocol.
Prepare the pretransformed library
strain (RFY231 + library plasmids)
4. Perform a large-scale transformation of EGY48 with
library DNA using the lithium
acetate method, starting with RFY231 bearing no other plasmids.
To prepare for transformation, grow RFY231 in YPD liquid medium.
Select library transformants on Glu/CM-Trp plates at 30oC
for 3 days.
5. Collect primary transformants by scraping plates,
washing yeast, and resuspending in 1 pellet vol glycerol solution
(65% (v/v) glycerol, 0.1M MGSO4, 25mM Tris, pH8.0). Freeze
0.2-1.0-ml aliquots at ?70 to ?80oC. The cells will be
stable for at least 1 year.
- Re-freezing a thawed aliquot will result in loss of viability.
Thus, many frozen aliquots should be made so that each thawed
aliquot can be discarded after use.
Prepare the pretransformed control
strain (RFY231 + pJG4-5)
6. Transform RFY231 grown in YPD liquid medium with the
empty library vector, pJG4-5, using the lithium
acetate method. Select transformants on Glu/CM-Trp plates at
30oC for 3 days.
7. Pick and combine three transformant colonies and use to
inoculate 30 ml of Glu/CM-Trp medium. Incubate 15-24 hours at
30oC to OD600 >3.
8. Centrifuge 5 min at 1000 to 1500 X g at room
temperature. Resuspend in 10 ml sterile water to wash cells.
9. Centrifuge 5 min at 1000 to 1500 X g at room
temperature. Resuspend in 1 pellet vol glycerol solution and freeze
100 µl aliquots at ?70 to ?80oC.
Determine plating efficiency of
pretransformed library and pretransformed control
strains.
10. After freezing (at least 1 hour) thaw an aliquot of
each pretransformed strain (from step 5 and step 9) at room
temperature. Make several serial dilutions in sterile water,
including aliquots diluted 105- 106-, and
107-fold. Plate 100 µl of each dilution on 100-mm
Glu/CM-Trp plates and incubate at 30oC for 2-3 days.
11. Count the colonies and determine the number of
colony-forming units (cfu) per aliquot of transformed yeast.
The plating efficiency for a typical library transformation
and for the control strain will be ~108 cfu/100
µl.
Mate the bait strain with the
pretransformed library strain and the pretransformed control
strain
In steps 12 through 20, an interactor hunt is conducted
concurrently with testing for LEU2 reporter activation by the
bait itself. For most baits this approach will be the quickest way to
isolate interactors. However, for some baits, such as those that have
a high transactivation potential, or those that affect yeast mating
or growth, steps 12 through 20 will serve as a pilot experiment to
determine the optimal parameters for a subsequent hunt.
12. Grow a 30 ml culture of the bait strain in
Glu/CM-Ura,-His liquid dropout medium to mid- to late-log phase
(OD600=1.0-2.0, or 2-4 x 107cells/ml)
A convenient way to grow the bait strain is to inoculate a 5
ml culture with ~ three colonies from a plate and grow it
overnight at 30oC with shaking. In the morning measure
the OD600, dilute into a 30 ml culture to a final
OD600=0.2, and grow at 30oC with shaking.
The culture should reach mid- to late-log phase before the end of
the day.
13. Centrifuge the culture 5 minutes at 1,000 - 1,500 X
g at room temperature to harvest cells. Resuspend the cell pellet
in sterile water to make a final volume of 1 ml. This should
correspond to ~1 x 109cells/ml.
14. Set up two matings. In one sterile eppendorf tube mix
200 µl of the bait strain with 200 µl of a thawed aliquot
of the pretransformed control strain from step 9. In a second
eppendorf tube mix 200 µl of the bait strain with approximately
108 cfu (~0.1 - 1 ml) of the pretransformed library strain
from step 5.
The library mating should be set up so that it contains a
~2-fold excess of bait strain cfu over pretransformed library
strain cfu. Because the bait strain was harvested in log phase,
most of the cells will be viable (i.e., cells/ml = ~cfu/ml), and
the number of cfu can be sufficiently estimated from optical
density (1 OD600=~2 X 107 cells/ml). Under
these conditions, ~10% of the cfu in the pretransformed library
strain will mate with the bait strain. Thus, a complete screen of
107 library transformants will require a single mating
with at least 108 cfu of the pretransformed library
strain and at least 2 X 108 cfu of the bait strain.
To screen more library transformants, set up additional
matings. The number of pretransformed library transformants to
screen depends on the size of the library and the number of
primary transformants obtained in step 5. If the size of the
library is larger than the number of transformants obtained in
step 5, the goal will be to screen all of the yeast transformants.
In this case, complete screening of the library will require
additional transformations of RFY231 and additional interactor
hunts. If the size of the library is smaller than the number of
transformants obtained in step 5, the goal will be to screen at
least a number of transformants equivalent to the size of the
library.
15. Centrifuge each cell mixture for 5 minutes at 1,000 -
1,500 X g, pour off media and resuspend cells in 200 µl
YPD. Plate each suspension onto a 100-mm YPD plate. Incubate 12-15
hours at 30oC.
16. Add ~ 1ml of Gal/Raff/CM-Ura,-His,-Trp to the lawns of
mated yeast on each plate. Mix the cells into the media using a
sterile applicator stick or glass rod.
17. Transfer each slurry of mated cells to a 500 ml flask
containing 100 ml of Gal/Raff/CM-Ura,-His,-Trp dropout medium.
Incubate with shaking 6 hrs at room temperature to induce the
GAL1 promoter which drives expression of the cDNA library.
18. Centrifuge the cell suspensions 5 minutes at 1,000 -
1,500 X g at room temperature to harvest the cells. Wash by
resuspending in 30 ml of sterile water and pelleting again. Resuspend
each pellet in 5 ml sterile water. Measure OD600 and, if
necessary, dilute to a final concentration of ~108
cells/ml.
This is a mixture of haploid cells that have not mated and
diploid cells. Under a microscope the two cell types can be
distinguished by size (diploids are ~1.7 times bigger than
haploids) and shape (diploids are slightly oblong and haploids are
spherical). Because diploids grow faster than haploids, this
mixture will contain about 10 ? 50% diploid cells. The actual
number of diploids will be determined by plating dilutions on
-Ura, -His, -Trp medium, which will not support the growth of the
parental haploids.
19. For each mating make a series of 1/10 dilutions, at
least 200 µl each, in sterile water to cover a
106-fold concentration range. Plate 100 µl from each
tube (undiluted, 10-1, 10-2, 10-3,
10-4, 10-5, and 10-6 dilution) on
100-mm Gal/Raff/CM-Ura,-His,-Trp,-Leu plates. Plate 100 µl from
tubes 10-4, 10-5, and 10-6 on 100-mm
Gal/Raff/CM-Ura,-His,-Trp plates. Incubate plates at 30oC.
Count the colonies on each plate after 2-5 days.
Selecting
interactors
20. For the mating with the pretransformed library, make an
additional 3 ml of a 10-1 dilution. Plate 100 µl of
the 10-1 dilution on twenty 100-mm
Gal/Raff/CM-Ura,-His,-Trp,-Leu plates, and 100 µl of the
undiluted cells on twenty 100-mm Gal/Raff/CM-Ura,-His,-Trp,-Leu
plates. Incubate at 30oC. Pick Leu+ colonies
after 2-5 days and characterize them as described in previous
protocols (See Part II D of the Finley,
1997)
The number of Leu+ colonies to pick to ensure that
all of the pretransformed library has been screened depends on the
transactivation potential of the bait protein itself. The
transactivation potential is expressed as the number of
Leu+ colonies that grow per cfu (Leu+/cfu) of the bait
strain mated with the control strain, as determined in step 19. It
can be calculated as the ratio of the number of colonies that grow
on Gal/Raff/CM-Ura,-His,-Trp,-Leu to the number of colonies that
grow on Gal/Raff/CM-Ura,-His,-Trp for a given dilution of the
mating between the bait strain and the control strain. A bait with
essentially no transactivation potential will produce less than
10-6 Leu+/cfu. For a bait to be useful in an
interactor hunt it should not transactivate more than
10-4 Leu+/cfu.
To screen all of the pretransformed library, it will be
necessary to pick a sufficient number of Leu+ colonies
in addition to background colonies produced by the transactivation
potential of the bait itself. Thus, the minimum number of
Leu+ colonies that should be picked at step 20 is given
by:
(transactivation potential, Leu+/cfu) x (# library
transformants screened).
For example, if 107 library transformants were
obtained in step 2 (and at least 108 cfu of these
transformants were mated with the bait strain in step 14, since
only ~10% will form diploids), and the transactivation potential
of the bait is 10-4 Leu+/cfu, then at least
1000 Leu+ colonies must be picked and characterized. In
other words, if the rarest interactor is present in the
pretransformed library at a frequency of 10-7, to find
it one needs to screen through at least 107 diploids
from a mating of the library strain. However, at least 1000 of
these 107 diploids would be expected to be
Leu+ due to the bait background if the transactivation
potential of the bait is 10-4. The true positives will
be distinguished from the bait background in the next step by the
galactose dependence of their Leu+ and lacZ+
phenotypes.
Back to Top
Literature Cited
Bendixen, C., Gangloff, S., Rothstein, R. 1994. A yeast
mating-selection scheme for detection of protein-protein
interactions. Nuclei Acids Research 22:1778-1779.
Brent, R., and Finley Jr., R.L. 1997. Understanding gene and
allele function with two-hybrid methods. Annual Review of Genetics
31: 663-704.
Estojak, J., Brent, R., and Golemis, E.A. 1995. Correlation of
two-hybrid affinity data with in vitro measurements. Mol. Cell.
Biol. 15, 5820-5829.
Fields, S., and Song, O. 1989. A novel genetic system to detect
protein-protein interaction. Nature 340, 245-246
Finley Jr., R.L. and Brent, R. 1994 Binary and ternary
interactions between Drosophila cell cycle regulators. Proc. Natl.
Acad. Sci. U.S.A. 91:12980-12984.
Finley Jr., R.L. and Brent, R. 1995. Interaction trap cloning
with yeast, in "DNA cloning-expression systems: a practical
approach"(ed. D. Glover and B.D. Hames) Oxford University Press,
Oxford, England, pp. 170-203.
Finley Jr., R.L. and Brent, R. 1997. Two-hybrid analysis of
genetic regulatory networks, in "The yeast two-hybrid system" (ed.
P.L. Bartel and S. Fields) Oxford University Press, Oxford,
England, pp. 197-214.
Finley Jr., R.L. 1997. Examining the function of proteins and
protein networks with the yeast two-hybrid system. April 1997
Methods Workshop, copyright, American Association for Cancer
Research (also here).
Golemis, E.A., Serebriiskii, I., Finley Jr., R.L, Kolonin,
M.G., Gyuris, J., and Brent, R. "Interaction Trap/Two-Hybrid
System to Identify Interacting Proteins" in Current Protocols in
Molecular Biology, Unit 20.1., and Current Protocols in Protein
Science, Unit 19.1, John Wiley and Sons, Inc., New York, NY,
1998.
Gyuris, J., Golemis, E., Chertkov, H., and Brent, R. 1993.
Cdi1, a human G1 and S phase protein phosphatase that associates
with Cdk2. Cell 75, 791-803.
Kolonin, M.G., and Finley Jr., R.L. Targeting cyclin-dependent
kinases in Drosophila with peptide aptamers. Proc. Natl. Acad.
Sci. U.S.A. 95: 14266-14271, 1998.
Sikorski, R.S. and Hieter, P. 1989. A system of shuttle vectors
and yeast host strains designed for efficient manipulation of DNA
in Saccharomyces cerevisiae. Genetics 122:19-27
|